Contents
Anatomy
The pituitary gland is a small gland that weighs around 500mg and is located beneath the frontal lobes of the brain in a small depression in the sphenoid bone known either as the sella turcica (which means Turkish saddle) or sometimes as the pituitary fossa. The term hypophysis can also be used for the pituitary and the adjective hypophyseal is derived from it.
The pituitary gland is composed of two components, the anterior and the posterior, which are also known as the adenohypophysis and the neurohypophysis respectively. These two parts of the gland have different functions and also have different embryological origins.
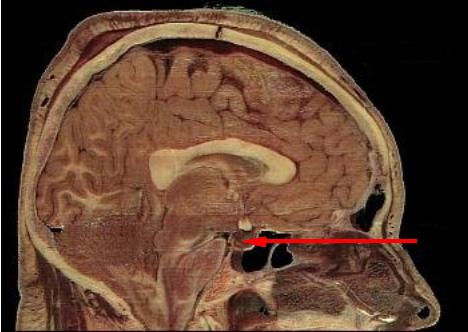
|
Sagittal section of a cadaver to show the pituitary (red arrow)
Image courtesy of Wikipedia
|
The anterior pituitary is derived from Rathke's pouch, which is an invagination of the posterior wall of the precursor of the pharynx that develops at 3 weeks and then grows upwards and backwards to reach its final position by 8 weeks, by which time it has lost contact with the pharynx.
The posterior pitituitary is an extension of the hypothalamus. The connection between the hypothalamus and the pituitary gland is known as the pituitary stalk or infundibulum.
The anterior pituitary gland possessses three parts. The pars distalis constitutes the bulk of the gland and executes most of the functions of the pituitary gland. The pars tuberalis forms a sheath around the pituitary stalk. The pars intermedia is located between the pars distalis and the posterior pituitary and minds its own business.
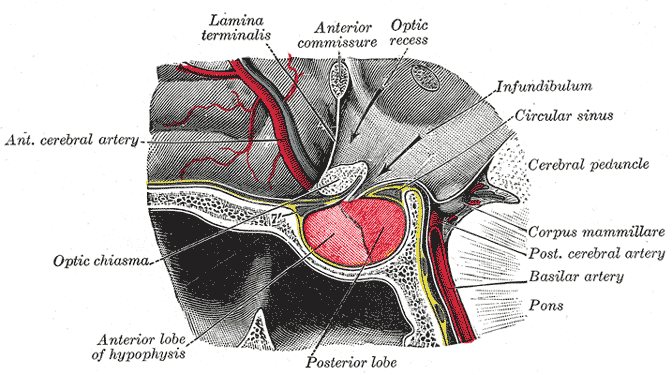
|
Sagittal view of the pituitary gland and its immediate neighbours. Anterior is on the left.
Diagram courtesy of Wikipedia
|
The adenohypophysis has a trabecular / glandular architecture. Staining techniques in the early days of histology delineated two main populations of glandular cells, chromophobes and chromophils. The chromophil cells were further classified into acidophil and basophil subtypes. Dilated, fenestrated capillaries are situated between the glandular cells. However, immunohistochemistry has largerly superseded the chromophobe / chromophil / acidophil / basophil classification.
The neurohypophysis is formed by the axon terminals of axons which arise from neuronal cell bodies that are located in the hypothalamus. The axon terminals contain numerous neurosecretory granules. Glial cells are also present.
The blood supply of the anterior part of the pituitary gland has an unusual arrangement. The purpose of this odd configuration is to allow communication between the hypothalamus and the pituitary gland. Rather than being supplied directly by the the superior hypophyseal arteries, the anterior pituitary receives blood via portal veins that drain a capillary plexus in the infundibulum. This capillary plexus is fed by the superior hypophyseal arteries. The hypothalamus releases hormones into this plexus in order to regulate the function of the anterior pituitary. This complicated arrangement is known as the hypothalamic hypophyseal portal system and is basically a way for the hypothalamus and the anterior pituitary gland to talk to each other by using hormones rather than direct innervation.
The posterior pituitary gland does not need to engage in such anatomical sleight of hand for its blood supply and just receives direct branches of the superior and inferior hypophyseal arteries.
The optic chiasm is located immediately above the pituitary fossa. This anatomical relation has important consequences for the clinical features of pituitary masses.
Anterior Pituitary Hormones
The role of the anterior pituitary explains why the pituitary is often known as the 'master gland', although the designation is a little inaccurate because the anterior pituitary is itself somewhat subservient to the hypothalamus. Nevertheless, one of the main roles of the anterior pituitary gland is to release hormones which control the function of several other endocrine glands. The adenohypophysis is stimulated to release these hormones in response to hormonal signals from the hypothalamus. The hormones that are generated by the glands which are controlled by the hypothalamus and pituitary exert negative feedback on the hypothalamus and pituitary to limit.
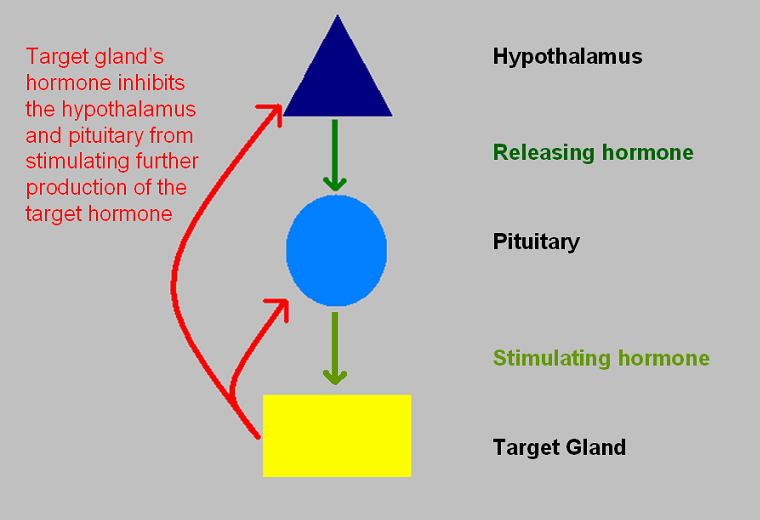
|
|
Illustration of the hypothalamic-pituitary-target gland axis and the negative feedback loops
|
The glands that are under the authority of the anterior pituitary are the thyroid, adrenal cortex (specifically glucocorticoid production) and the gonads.
The production of thryoxine by the thyroid gland is stimulated by the release of thyroid stimulating hormone (TSH) by the adenohypophysis. Thyroid stimulating hormone is a glycoprotein that has an alpha and beta subunit. The alpha subunit is the same as that employed by some other anterior pituitary hormone whereas the beta subunit is individual to TSH and enables it to exert its specific function. The release of TSH is in turn governed by the secretion of thyroid releasing hormone (TRH) by the hypothalamus.
The synthesis of glucocorticoids by the
adrenal cortex is regulated by the production of adrenocorticotrophic hormone (ACTH) by the anterior pituitary. ACTH is a peptide hormone which possesses 39 amino acids and is produced by the cleavage of the larger precursor peptide, preproopiomelanocortin. The hypothalamus stimulates the anterior pituitary to generate ACTH by using corticotropin releasing hormone (CRF).
The gonads receive dual control from the anterior pituitary in the form of follicle stimulating hormone (FSH) and luteinising hormone (LH). Both FSH and LH are glycoproteins that have alpha and beta subunits. The alpha subunit is the same as that employed by TSH (and also by human chorionic gonadotropin). The levels of FSH and LH are regulated by the production of gonadotropin releasing hormone by the hypothalamus (GNRH or GnRH). The hypothalamic-pituitary-ovarian axis exhibits feedback patterns which are more complicated than the simple negative feedback loop discussed above.
As well as the hormones that it uses to control other glands, the anterior pituitary releases two hormones that act directly on end organs. These two hormones are prolactin and growth hormone. The cells of the anterior pituitary which synthesise these two hormones are of the acidophil type while the quartet of gland-stimulating hormones are made by basophilic cells. Prolactin and growth hormone also share the property of being under dual control by the hypothalamus. Prolactin release is inhibited by dopamine but can also be stimulated by TRH (the former is probably the dominant method of control). Growth hormone release is stimulated by the appropriately named growth hormone releasing hormone (GHRH) but can be inhibited by somatostatin.
Posterior Pituitary Hormones
The posterior pituitary gland is a rather different kettle of fish from the anterior pituitary. In effect it is simply a collection of axon terminals of hypothalamic neurones which are gathered into a place where they can release the hormones they synthesise into the capillaries of the pituitary gland, from where the hormones will be disseminated throughout the body.
Two posterior pituitary hormones exist, antidiuretic hormone (ADH) and oxytocin.
Antidiuretic Hormone
Antidiuretic hormone is a small peptide hormone that is sometimes known as vasopression. It is only nine amino acids long and has a sequence of Cys-Tyr-Phe-Gln-Asn-Cys-Pro-Arg-Gly-NH2. It is manufactured in the supra-optic and paraventricular nuclei of the hypothalamus.
The main role of ADH is to conserve water. Unlike
aldosterone it accomplishes this without producing a parallel retention of sodium; ADH promotes the retention of pure water and it is thus the key regulator of plasma osmolality, as well as being vital in the maintenance of total body water.
ADH is released in response to an increase in blood osmolality or a fall in blood volume; the former is the more powerful stimulus. The neurones that make ADH have osmoreceptor abilities, although they are not the only osmoreceptors in the body and receive input from these other osmoreceptors, as well as various blood pressure sensors.
ADH acts on the kidney to promote the retention of water. This effect is accomplished by instructing the collecting ducts of the nephron to express greater numbers of the water channel, acquaporin 2. The acquaporin channel permits water to move out of the lumen of the nephron back into the blood. The second messenger system that mediates the increased expression of acquaporin 2 channels is cyclic AMP. The increase in the water reabsorption by the kidney causes the urine to become more concentrated.
The efficacy of the increase in the number of water channels is enhanced by the stimulation of the countercurrent mechanism by ADH. The countercurrent mechanism is a nifty manoeuvre which is performed by the loop of Henle of the nephron to generate an osmotic gradient that can be exploited by the nephron, including the collecting ducts.
The participation of ADH in preserving body water does not stop at the nephron. ADH also induces a sense of thirst and therefore encourages drinking, which is a good way to replenish lost water.
In keeping with the role of a low blood pressure / low blood volume in triggering the release of ADH, antidiuretic hormone also possesses vasoconstrictor powers. Under normal circumstances the contribution of ADH to vasoconstriction is rather limited, but in the situation of a large haemorrhage the resulting rise in ADH and the vasoconstriction it induces can help to assist the attempts by the body to preserve blood pressure.
Antidiuretic has also been suggested to be involved in some aspects of the function of the central nervous system, including mating behaviour and mating bonding.
Oxytocin
Oxytocin is also a nonapeptide that is synthesised by the supraoptic and paraventricular nuclei of the hypothalamus. Its sequence is Cys-Tyr-Ile-Gln-Asn-Cys-Pro-Leu-Gly-NH2 and thus it differs by only two amino acids from ADH. However, this difference is sufficient to impart very different actions to oxytocin.
Oxytocin was initially considered to be a hormone that was only of real importance in females. It triggers the 'letdown' reflex in breastfeeding which allows milk to be secreted via the nipple. The stimulation of the nipple by the suckling baby is important for evoking oxytocin release by the hypothalamus-neurohypophysis.
Contraction of the uterus during labour is stimulated by oxytocin (among other factors). The contractions themselves stimulate the release of oxytocin and thus a positive feedback loop is generated.
More recent research has concentrated on the role of oxytocin in sexual behaviour and this has begun to disclose the functions of oxytocin in males, as well as further properties in females. A surge of oyxtocin release occurs at orgasm and is believed to be involved in bonding between partners, including inducing feelings of contentment, calmness and security. It has also been suggested that the burst of oxytocin activity induces sleep in males.
